






- ダルマガエルの危機的状況(松本盆地:10年間で悪化 、伊那盆地:依然として厳しい状況)
- トウキョウダルマガエル分布域(長野盆地)にトノサマガエルの進入を初確認
- ナゴヤダルマガエル分布域(伊那盆地)からトウキョウダルマガエルDNA検出
本研究のポイント
● 信州はトノサマガエル・ダルマガエルの分布が重なる重要地域(ともに絶滅危惧種としてレッドリストに登載)
● ダルマガエルは以下の2亜種に区分(東日本:トウキョウダルマガエル、西日本:ナゴヤダルマガエル)
トウキョウダルマガエル(松本盆地・長野盆地など)
ナゴヤダルマガエル(伊那盆地)
● 1980年代以降、両種の交雑進行が危惧され、2010年に遺伝子解析による実態調査を実施
● 前回調査から10年後(2020年)に遺伝子解析の再調査を実施し、その動態を英国専門誌に公表
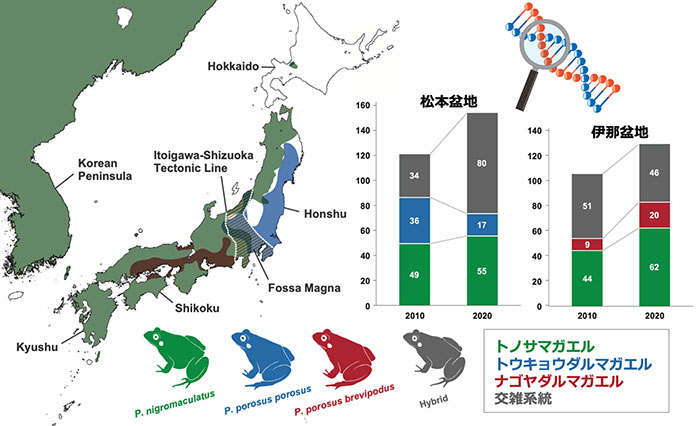
● 松本盆地では10年間で両種の純系個体の割合が低下し、交雑個体の割合が増加
● 伊那盆地では、10年間で両種の純系個体の割合がやや増加したものの、純系のナゴヤダルマガエルは依然として少ない状況(2010年には純系個体だけで構成されていた個体群でも交雑系統が確認された)
● ナゴヤダルマガエルが生息する伊那盆地内で、トウキョウダルマガエル型のミトコンドリアDNAをもつ個体が確認された(トノサマガエルとの交雑系統を経由して、DNAだけが運ばれた?)
● ダルマガエルは以下の2亜種に区分(東日本:トウキョウダルマガエル、西日本:ナゴヤダルマガエル)
トウキョウダルマガエル(松本盆地・長野盆地など)
ナゴヤダルマガエル(伊那盆地)
● 1980年代以降、両種の交雑進行が危惧され、2010年に遺伝子解析による実態調査を実施
● 前回調査から10年後(2020年)に遺伝子解析の再調査を実施し、その動態を英国専門誌に公表
● 松本盆地では10年間で両種の純系個体の割合が低下し、交雑個体の割合が増加
● 伊那盆地では、10年間で両種の純系個体の割合がやや増加したものの、純系のナゴヤダルマガエルは依然として少ない状況(2010年には純系個体だけで構成されていた個体群でも交雑系統が確認された)
● ナゴヤダルマガエルが生息する伊那盆地内で、トウキョウダルマガエル型のミトコンドリアDNAをもつ個体が確認された(トノサマガエルとの交雑系統を経由して、DNAだけが運ばれた?)
研究の背景
信州大学学術研究院・理学系の東城幸治教授と研究室の大学院生(総合理工学研究科 理学専攻の重田将之介氏)、東城研究室出身の小巻翔平博士(岩手医科大学)と鈴木智也博士(広島修道大学)で構成される研究グループは、互いに近縁な姉妹種で、以下に示すように環境省や自治体レベルでの絶滅危惧種として扱われているトノサマガエルとダルマガエルの生息域や遺伝構造、それらの動態に関する長期的モニタリング結果を取りまとめ、この度、10年間(2010−2020年)での動態についてまとめた論文を、英国生態学会が刊行する進化生態学の専門誌 Ecology and Evolution に公表した。
トノサマガエル Pelophylax nigromaculatus 環境省:準絶滅危惧、長野県:準絶滅危惧
トウキョウダルマガエル Pelophylax porosus porosus 環境省:準絶滅危惧、長野県:絶滅危惧II類
ナゴヤダルマガエル Pelophylax porosus brevipodus 環境省:絶滅危惧IB、長野県:絶滅危惧I類
信州は、これらのPelophylax属カエルの分布域が、互いに接したり、重なり合う興味深い地域であるが、これらのカエル本来の生息地である河川氾濫原の湿地的環境が激減してきたことを受け、現在は、いずれも絶滅危惧種として扱われている。氾濫原に代わり、これらのカエル類の代替生息地として利用されてきた水田についても、国の減反政策により、宅地や畑地への農地転換や、圃場整備事業、農業形態の画一化(代掻きや田植え、中干しの同調化や長期化)など、ハビタット(生息場)自体の減少による個体密度の低下や、繁殖時期の季節的な種差が縮小することによる種間交雑の進行が危惧されている。
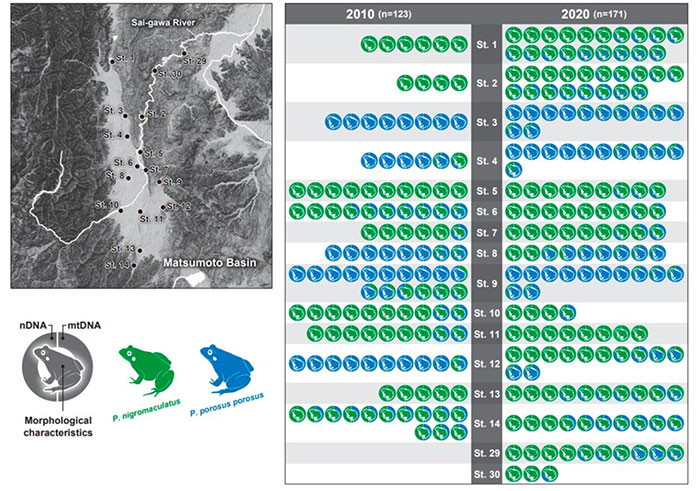
長野県内の松本盆地や伊那盆地では、1980年代から両種の分布域や種間交雑に関する調査研究が実施されてきた。2010年には、本研究グループの小巻博士が中心となり、松本・伊那両盆地の広域を対象に、松本盆地内14地点から119個体、伊那盆地内12地点から104個体のPelophylax属カエルを採取し、形態形質の評価と遺伝子解析(核DNA、ミトコンドリアDNA)を伴う、詳細かつ大々的な分布調査や交雑状況の調査研究を実施した(Komaki et al., 2012)。この先行研究では、1980年代から2010年までの30年間にわたる分布域の動態や、2010年当時の種間交雑の状況を報告している。
本研究は、この2010年に実施した先行研究から10年が経過した2020年に同様の調査研究を実施し、2010年から2020年までの10年間での分布動態や種間交雑の推移を評価したものである。なお、今回の調査研究においては、重田氏が中心となり、現地調査や遺伝子解析を実施したものである。
トノサマガエル Pelophylax nigromaculatus 環境省:準絶滅危惧、長野県:準絶滅危惧
トウキョウダルマガエル Pelophylax porosus porosus 環境省:準絶滅危惧、長野県:絶滅危惧II類
ナゴヤダルマガエル Pelophylax porosus brevipodus 環境省:絶滅危惧IB、長野県:絶滅危惧I類
信州は、これらのPelophylax属カエルの分布域が、互いに接したり、重なり合う興味深い地域であるが、これらのカエル本来の生息地である河川氾濫原の湿地的環境が激減してきたことを受け、現在は、いずれも絶滅危惧種として扱われている。氾濫原に代わり、これらのカエル類の代替生息地として利用されてきた水田についても、国の減反政策により、宅地や畑地への農地転換や、圃場整備事業、農業形態の画一化(代掻きや田植え、中干しの同調化や長期化)など、ハビタット(生息場)自体の減少による個体密度の低下や、繁殖時期の季節的な種差が縮小することによる種間交雑の進行が危惧されている。
長野県内の松本盆地や伊那盆地では、1980年代から両種の分布域や種間交雑に関する調査研究が実施されてきた。2010年には、本研究グループの小巻博士が中心となり、松本・伊那両盆地の広域を対象に、松本盆地内14地点から119個体、伊那盆地内12地点から104個体のPelophylax属カエルを採取し、形態形質の評価と遺伝子解析(核DNA、ミトコンドリアDNA)を伴う、詳細かつ大々的な分布調査や交雑状況の調査研究を実施した(Komaki et al., 2012)。この先行研究では、1980年代から2010年までの30年間にわたる分布域の動態や、2010年当時の種間交雑の状況を報告している。
本研究は、この2010年に実施した先行研究から10年が経過した2020年に同様の調査研究を実施し、2010年から2020年までの10年間での分布動態や種間交雑の推移を評価したものである。なお、今回の調査研究においては、重田氏が中心となり、現地調査や遺伝子解析を実施したものである。
研究方法
先行研究(Komaki et al., 2012)による全ての調査地点を対象とし、さらに松本盆地の下流域にあたり、長野盆地へと繋がる犀川流域にも新たに2つの調査地点を設定した。また、伊那盆地の南部にも2つの調査地点を追加した。松本盆地(および犀川流域)内の16地点から165個体、伊那盆地内の14地点から129個体のPelophylax属カエルを採取し、形態形質の評価と遺伝子解析(核DNA、ミトコンドリアDNA)を実施した。
形態形質については、両種の識別において有用な形質を評価するとともに、全ての解析個体について、体背面と体側面からの写真を撮影した。遺伝子解析については、核DNAのチロシナーゼ領域(747塩基)とミトコンドリアDNAのシトクロムb遺伝子(610塩基)を解析した。形態形質、核DNA、ミトコンドリアDNAの解析結果に齟齬がみられた場合には、当該個体を交雑系統として評価した。種間交雑により生まれた雑種第一代(F1)のオスは不稔となる一方で、F1メスは妊性があるため、いずれかの親種(あるいは交雑系統)のオスとの間での戻し交雑が可能となる。こうした戻し交雑を繰り返した交雑系統に関しては、今回の判定法では「交雑」系統ではなく、「純系」として評価されてしまう可能性があり、本研究の結果は、交雑系統の割合を過小評価している可能性がある。ただし、核DNAとミトコンドリアDNAによる種判別により「交雑」系統であると評価された個体については、間違いなく純系ではなく、過去に種間交雑を経験した系統であることから、交雑系統の割合を過大評価することはない。すなわち、本研究における目的の一つである、種間交雑の進行状況の把握や長期的なモニタリングにおいては、一定の評価が可能である。
形態形質については、両種の識別において有用な形質を評価するとともに、全ての解析個体について、体背面と体側面からの写真を撮影した。遺伝子解析については、核DNAのチロシナーゼ領域(747塩基)とミトコンドリアDNAのシトクロムb遺伝子(610塩基)を解析した。形態形質、核DNA、ミトコンドリアDNAの解析結果に齟齬がみられた場合には、当該個体を交雑系統として評価した。種間交雑により生まれた雑種第一代(F1)のオスは不稔となる一方で、F1メスは妊性があるため、いずれかの親種(あるいは交雑系統)のオスとの間での戻し交雑が可能となる。こうした戻し交雑を繰り返した交雑系統に関しては、今回の判定法では「交雑」系統ではなく、「純系」として評価されてしまう可能性があり、本研究の結果は、交雑系統の割合を過小評価している可能性がある。ただし、核DNAとミトコンドリアDNAによる種判別により「交雑」系統であると評価された個体については、間違いなく純系ではなく、過去に種間交雑を経験した系統であることから、交雑系統の割合を過大評価することはない。すなわち、本研究における目的の一つである、種間交雑の進行状況の把握や長期的なモニタリングにおいては、一定の評価が可能である。
結果と考察
松本盆地におけるトノサマガエル・トウキョウダルマガエルの分布と種間交雑状況
伊那盆地におけるトノサマガエル・トウキョウダルマガエルの分布と種間交雑状況

松本盆地・伊那盆地における分布動態、および種間交雑動態
トノサマガエル、トウキョウダルマガエル、ナゴヤダルマガエルそれぞれの純系、交雑系統の割合の10年間(2010 年−2020 年)での変化については、冒頭の棒グラフに示した通りである。各調査地点内の個体レベルでの形態形質や遺伝子解析による純系/交雑系統の評価については、前頁に松本盆地(上段図)・伊那盆地(下段図)それぞれに分けて示した。
松本盆地では、トノサマガエルとトウキョウダルマガエルそれぞれの純系個体の割合が減少し、交雑個体が増加する傾向が確認された。また、2010 年の調査研究時にはトノサマガエル純系やトウキョウダルマガエル純系だけで構成されていた個体群についても、2020 年には種間交雑系統が含まれるように変化していた。
伊那盆地では、絶滅が危惧されるナゴヤダルマガエルの純系個体の割合が増加傾向を示したものの、依然として純系は少なく、またナゴヤダルマガエル純系だけで構成される個体群は、2010 年、2020 年の調査のいずれにおいても認められなかった。トノサマガエルの純系個体だけで構成される個体群は2010 年調査では2個体群、2020 年調査で3個体群が確認された。
松本盆地では、トノサマガエルとトウキョウダルマガエルそれぞれの純系個体の割合が減少し、交雑個体が増加する傾向が確認された。また、2010 年の調査研究時にはトノサマガエル純系やトウキョウダルマガエル純系だけで構成されていた個体群についても、2020 年には種間交雑系統が含まれるように変化していた。
伊那盆地では、絶滅が危惧されるナゴヤダルマガエルの純系個体の割合が増加傾向を示したものの、依然として純系は少なく、またナゴヤダルマガエル純系だけで構成される個体群は、2010 年、2020 年の調査のいずれにおいても認められなかった。トノサマガエルの純系個体だけで構成される個体群は2010 年調査では2個体群、2020 年調査で3個体群が確認された。
伊那盆地からトウキョウダルマガエル型DNA の検出
また、伊那盆地のダルマガエルは、ナゴヤダルマガエル亜種だけが生息するとされ、トウキョウダルマガエル亜種が生息する松本盆地との間に、これら両亜種の分布境界が存在するとされるが、この研究では、伊那盆地内の3地点(地点No. 21, 27, 28)において、トウキョウダルマガエル型のDNA 配列が検出された。
かつて、これら両亜種の分布域が接した時期があり、その際に亜種間での遺伝子浸透が生じた名残りであるのかもしれないが、そうであれば伊那盆地の北部(松本盆地近く)においてこうした個体が検出されやすいはずであり、逆に松本盆地内でナゴヤダルマガエル型のDNA が検出されてもよさそうであるが、そうした傾向はみられない。地点No. 21 は伊那盆地の北部であるが、No. 27, 28 は最南部であることも不可解である。
ダルマガエルよりも大型で、かつ移動力も大きなトノサマガエルとトウキョウダルマガエル間での種間交雑を経験した交雑系統による分散が起因している可能性も考えられる。実際に、トノサマガエル・オスとトウキョウダルマガエル・メス間での種間交雑が多く(逆に伊那盆地では、ナゴヤダルマガエル・オスとトノサマガエル・メスの組み合わせでの種間交雑が多い)、そうした交雑個体のメスがトノサマガエルとの戻し交雑を繰り返しながら分散した結果であるかもしれず、この課題については、今後、ゲノムワイドなSNPs解析などを通して詳細に検討したい。
かつて、これら両亜種の分布域が接した時期があり、その際に亜種間での遺伝子浸透が生じた名残りであるのかもしれないが、そうであれば伊那盆地の北部(松本盆地近く)においてこうした個体が検出されやすいはずであり、逆に松本盆地内でナゴヤダルマガエル型のDNA が検出されてもよさそうであるが、そうした傾向はみられない。地点No. 21 は伊那盆地の北部であるが、No. 27, 28 は最南部であることも不可解である。
ダルマガエルよりも大型で、かつ移動力も大きなトノサマガエルとトウキョウダルマガエル間での種間交雑を経験した交雑系統による分散が起因している可能性も考えられる。実際に、トノサマガエル・オスとトウキョウダルマガエル・メス間での種間交雑が多く(逆に伊那盆地では、ナゴヤダルマガエル・オスとトノサマガエル・メスの組み合わせでの種間交雑が多い)、そうした交雑個体のメスがトノサマガエルとの戻し交雑を繰り返しながら分散した結果であるかもしれず、この課題については、今後、ゲノムワイドなSNPs解析などを通して詳細に検討したい。
松本盆地より下流域: 犀川流域、および長野盆地内での分布動態、種間交雑動態
2010 年の先行研究の段階で、松本盆地から犀川沿いに下った地点(No. 2 の生坂村)において、純系のトノサマガエルが確認されていた。元来、犀川流域や、より下流域にあたる長野盆地はトウキョウダルマガエルだけが生息する地域と考えられてきたため、トノサマガエルや交雑系統のより下流域への分布拡大が危惧されたことで、2020 年の調査では、さらに下流域の2地点(No. 29, 30, それぞれ長野市信州新町平水内と長野市大岡甲)を追加したところ、これら2地点からは純系のトノサマガエル(6/13 個体)と交雑個体(7/13個体)だけが検出され、純系のトウキョウダルマガエルは確認されなかった。
この結果を受けて、今回、論文発表した内容には含まれていないものの、2024 年に長野盆地内で同様の調査を実施したところ、長野盆地内の2地点(長野市安茂里、長野市真島町川合)からトノサマガエル純系個体と交雑個体が確認された。長野市安茂里から採取された7個体のPelophylax 属カエルのうち、5個体は純系トノサマガエルであり、残る2個体は交雑系統であった。また、長野市真島町川合で採取された4個体は全て交雑系統であった。このほか、長野盆地内広域を対象にした調査解析を実施したが、2024 年段階でトノサマガエル型DNA が検出されたのは、犀川沿いの長野盆地の最上端部だけであった。ただし、犀川沿いに流下する方向での分散は起こりやすく、千曲川との合流地点のすぐ近くにまでトノサマガエル系統が分布している状況から、今後、注意深くモニタリングしていく必要がある。
この結果を受けて、今回、論文発表した内容には含まれていないものの、2024 年に長野盆地内で同様の調査を実施したところ、長野盆地内の2地点(長野市安茂里、長野市真島町川合)からトノサマガエル純系個体と交雑個体が確認された。長野市安茂里から採取された7個体のPelophylax 属カエルのうち、5個体は純系トノサマガエルであり、残る2個体は交雑系統であった。また、長野市真島町川合で採取された4個体は全て交雑系統であった。このほか、長野盆地内広域を対象にした調査解析を実施したが、2024 年段階でトノサマガエル型DNA が検出されたのは、犀川沿いの長野盆地の最上端部だけであった。ただし、犀川沿いに流下する方向での分散は起こりやすく、千曲川との合流地点のすぐ近くにまでトノサマガエル系統が分布している状況から、今後、注意深くモニタリングしていく必要がある。
今後の課題
2010 年、2020 年に引き続き、2030 年にも10 年周期での同地域内での同様の調査解析を実施し、トノサマガエルとダルマガエルの長期的な分布動態、交雑動態をモニタリングする予定である。また、今回新たに、長野盆地内へのトノサマガエル(あるいは交雑系統)の進入(分布拡大)が確認されたことを受け、千曲川流域を中心とする長野盆地内広域的な調査モニタリングが重要となる。
これまでの研究結果からは、トノサマガエルの河川流路に沿った上流域から下流域への分布域拡大が明らかになっており、千曲川に犀川が合流する流域まで交雑系統も含むトノサマガエルの分布拡大が新たに確認されたことから、今後は、千曲川本流を流下する方向での分散が危惧される。しかし、千曲川・犀川合流地点より上流側においても、河床勾配がかなり緩やかであることから、長野市内から千曲市内、坂城町や上田市側へと上流側へ分布拡大する可能性も考えられる。今後は、長野盆地だけでなく、より対象エリアを上流側へも拡大した調査研究も必要である。
これまでの研究結果からは、トノサマガエルの河川流路に沿った上流域から下流域への分布域拡大が明らかになっており、千曲川に犀川が合流する流域まで交雑系統も含むトノサマガエルの分布拡大が新たに確認されたことから、今後は、千曲川本流を流下する方向での分散が危惧される。しかし、千曲川・犀川合流地点より上流側においても、河床勾配がかなり緩やかであることから、長野市内から千曲市内、坂城町や上田市側へと上流側へ分布拡大する可能性も考えられる。今後は、長野盆地だけでなく、より対象エリアを上流側へも拡大した調査研究も必要である。
本研究の学術的な意義
ここまで、トノサマガエル・ダルマガエルがともに絶滅危惧種であることや、その純系個体が減少していること、一方で、交雑系統も含むトノサマガエルの分布域が拡大している現状を中心に記述してきた。これらは主に保全生態学や保全遺伝学的な視点からの議論である。一方、この研究の生物学的な意義としては、近縁な姉妹種の二次的接触という興味深い現象に焦点を当てた面にも触れておく必要がある。
生物学における種内個体群の分化や種分化は、遺伝的多様性や種の多様性の創出に繋がる最重要課題の一つである。地理・地形や地史などの影響による生息域の分断は、それぞれの系統や種の形質に特有の変異をもたらし、系統や種の分化・多様化を促進する。こうした形質の分化により生殖的隔離が促進され、完全な生殖的隔離が成立した場合には種分化の成立となる。実際には、ひとたび分化した系統や種が再び接触することも多く、こうした二次的接触による様々な興味深い事例が報告されている。特に、ゲノムワイドな遺伝構造の解析から、遺伝子流動の実態(強度や方向性など)が議論しやすくなった昨今、注目が集まりつつある。生殖的隔離が不十分である場合における二次的接触や、種分化が成立した後の二次的接触による交雑がさまざまな結果をもたらすことが報告されている。独自の変異を遂げた系統間の交雑が新たな遺伝系統を創出し、適応的な遺伝的変異の源となる可能性もあれば、生態学的適応度の低下に起因する不適合交雑を介し、種間の区別を強化することなどが議論されている。
こうした二次的接触帯における多様かつ複雑な現象の理解は、種の維持や多様化メカニズムの理解を深めることとなり、生物進化学・多様性生物学的観点においても極めて重要かつ興味深いことである。本研究における、信州を舞台としたトノサマガエル・ダルマガエルの進化生態学的研究の成果は、こうした観点においても極めて興味深い内容を含む研究と言える。
生物学における種内個体群の分化や種分化は、遺伝的多様性や種の多様性の創出に繋がる最重要課題の一つである。地理・地形や地史などの影響による生息域の分断は、それぞれの系統や種の形質に特有の変異をもたらし、系統や種の分化・多様化を促進する。こうした形質の分化により生殖的隔離が促進され、完全な生殖的隔離が成立した場合には種分化の成立となる。実際には、ひとたび分化した系統や種が再び接触することも多く、こうした二次的接触による様々な興味深い事例が報告されている。特に、ゲノムワイドな遺伝構造の解析から、遺伝子流動の実態(強度や方向性など)が議論しやすくなった昨今、注目が集まりつつある。生殖的隔離が不十分である場合における二次的接触や、種分化が成立した後の二次的接触による交雑がさまざまな結果をもたらすことが報告されている。独自の変異を遂げた系統間の交雑が新たな遺伝系統を創出し、適応的な遺伝的変異の源となる可能性もあれば、生態学的適応度の低下に起因する不適合交雑を介し、種間の区別を強化することなどが議論されている。
こうした二次的接触帯における多様かつ複雑な現象の理解は、種の維持や多様化メカニズムの理解を深めることとなり、生物進化学・多様性生物学的観点においても極めて重要かつ興味深いことである。本研究における、信州を舞台としたトノサマガエル・ダルマガエルの進化生態学的研究の成果は、こうした観点においても極めて興味深い内容を含む研究と言える。
論文タイトル
Changes over a 10-year period in the distribution ranges and genetic hybridization of three Pelophylax pond frogs in Central Japan
論文へのリンク(DOI)
著者
信州大学(大学院 総合理工学研究科 修士2 年) 重田 将之介
広島修道大学 人間環境学部 鈴木智也
岩手医科大学 いわて東北メディカル・メガバンク機構生体情報解析部門 小巻 翔平
信州大学(学術研究院 理学系, 山岳科学研究所) 東城 幸治(責任著者)
広島修道大学 人間環境学部 鈴木智也
岩手医科大学 いわて東北メディカル・メガバンク機構生体情報解析部門 小巻 翔平
信州大学(学術研究院 理学系, 山岳科学研究所) 東城 幸治(責任著者)
掲載誌
Ecology and Evolution 15: e71856(米国・進化生態学分野の専門誌;2025 年8 月7 日公開)